He colgado en la sección de apuntes y de presentaciones todos los documentos que cubren el temario de Biología de Segundo de Bachillerato posteriormente a la implantación de la LOMLOE.
Se basan en el temario de PAU vigente para el Principado de Asturias.
Durante el último año he elaborado y publicado en mi canal de YouTube una serie de vídeos para la preparación de la EBAU de Biología. Se trata de vídeos con la resolución de ejercicios correspondientes a la EBAU de Biología del Principado de Asturias.
Del primer bloque, hay un vídeo sobre preguntas relacionadas con el ciclo del agua:
Del bloque de Biología Celular hay un vídeo sobre esquemas de células:
Y otro vídeo sobre esquemas de metabolismo:
También podemos encontrar dos vídeos con problemas de genética de la EBAU resueltos:
El nitrógeno es un elemento esencial para los seres vivos, tanto en cuanto forma parte de las moléculas orgánicas que componen las proteínas (aminoácidos) y los ácidos nucléicos (bases nitrogenadas). Se trata de un elemento químico muy abundante en la atmósfera, pero su fijación en forma de nitrógeno orgánico es muy compleja, pues se encuentra en forma de N2 que es químicamente inerte y recae de forma exclusiva en los microorganismos.
La mayor parte de los organismos, por lo tanto, dependen de otras formas de nitrógeno para su utilización, tales como el amoniaco, nitrato o compuestos orgánicos ricos en nitrógeno. La mayor parte de los flujos de nitrógeno, por lo tanto, se llevan a cabo entre estas otras formas químicas, que son más accesibles a los seres vivos.
La Fijación del nitrógeno atmosférico es la transformación del nitrógeno atmosférico en otras formas de nitrógeno y depende de una serie de bacterias denominadas diazótrofas. Mediante un complejo enzimático denominado Nitrogenasa transforman el nitrógeno atmosférico, es decir, N2, en formas de nitrógeno accesibles para los seres vivos, principalemente amoniaco, es decir, NH3.
Entre las bacterias fijadoras del nitrógeno se distinguen dos grandes grupos, las bacterias de vida libre y las bacterias que se encuentran en simbiosis con algún otro organismo. Entre las bacterias de vida libre destacan algunos grupos de gramnegativos, como las bacterias del género Azotobacter, así como algunas cianobacterias. Las simbióticas viven asociadas a algún ser vivo, fundamentalmente plantas. Les ceden los compuestos nitrogenados derivados del amoniaco y a cambio reciben materia orgánica fabricada por el organismo al que se asocian. Un ejemplo son las bacterias del género Rhizobium que viven asociados a las raíces de plantas, o cianobacterias como la Anabeana que crecen asociadas a helechos acuáticos.
Una vez fijado, el nitrógeno es captado por los organismos, principalmente los productores, que lo incorporan en forma de ion nitrato (NO3-) o amoniaco (NH3). Este proceso se denomina Asimilación.
El nitrógeno contenido en las moléculas orgánicas puede tender a acumularse en algunos organismos que no tienen capacidad de oxidar nitrógeno, siendo transformado para formar sustancias de desecho de las que tendrá que deshacerse. Los sistemas de eliminación de los excesos de nitrógeno varían ligeramente entre distintos seres vivos, pudiendo transformarse en amoniaco, urea, ácido úrico o incluso guanina.
Estos restos nitrogenados, junto con proteínas aminoácidos liberados o procedentes de otros organismos son utilizados por ciertos organismos que acabarán transformando el nitrógeno en la forma más simple, el amoniaco (NH3). Este proceso se denomina Amonificación. Y es llevado a cabo por bacterias amonificantes.
Las bacterias que fijan el nitrógeno atmosférico también llevan a cabo procesos de amonificación para transformarlo en amoniaco.
Algunas bacterias usan el amoniaco para generar iones nitrato, obteniendo energía en el proceso. El proceso se denomina Nitrificación y es llevado a cabo por bacterias quimiolitótrofas de los géneros Nitrosomonas y Nitrobacter. El proceso de transformación requiere oxígeno, por lo que estas bacterias son aeróbicas.
El nitrógeno transformado por las bacterias nitrificantes en nitratos puede ser incorporado a algunas plantas mediante procesos de asimilación (similares a los procesos de asimilación que tienen lugar cuando una bacteria fijadora le cede nitrógeno a la planta con la que se encuentra en simbiosis).
Para mantener el equilibrio, debe existir algún sistema de transformar el nitrógeno fijado en forma de amoniaco o de nitrato en nitrógeno molecular gaseoso (N2) de nuevo (de no ser así, el nitrógeno atmosférico tendería a disminuir). Esto tiene lugar mediante un proceso que conocemos como Desnitrificación. Y es llevado a cabo por ciertas bacterias heterótrofas, que aprovechan el proceso para obtener energía, como por ejemplo Pseudomonas fluorescens. Este grupo de bacterias usan amonio y nitritos como aceptores finales de electrones en sustitución del oxígeno (es decir, se trata de reacciones anaeróbicas) y se denominan genéricamente bacterias desnitrificantes. Y el conjunto de reacciones catabólicas que conducen a la transformación de amoniaco y nitrito en nitrógeno molecular se denomina oxidación anaerobia del ion amonio o Anammox.
He comenzado a redactar un nuevo lote de apuntes sobre Biología y Geología. Se trata de temas individuales de Biología y Geología con un nivel calculado para primero de bachillerato.
Se corresponden con las presentaciones que ya estaban disponibles de cursos anteriores, pero en este caso se trata de texto con imágenes que puede ayudar a complementar los contenidos desarrollados de forma esquemática en las presentaciones.
Como siempre, se trata de textos en los que se han usado bien materiales de código abierto, bien materiales propios, por lo que pueden ser usados y descargados sin ningún tipo de problema.
Espero que sean de utilidad.
De momento se han incorporado los temas correspondientes al primer trimestre, pero se irán incorporando sucesivos temas conforme vaya avanzando el curso.
En ocasiones, la expresión de un gen interactúa o condiciona la expresión de otros genes, que pueden estar relacionados con distintos alelos. Es decir, puede suceder que la expresión de un gen interactúe con otros genes y provoque que estos no se expresen o que la expresión de los mismos sea diferente a la esperada o no se manifieste con el fenotipo esperado.
Esta interacción entre genes puede hacer que las proporciones de los descendientes no sea la esperada o no se corresponda con la descendencia prevista según las leyes mendelianas de la herencia.
Estas variaciones de las proporciones esperadas en un alelo concreto y derivadas de la expresión de genes relacionados con alelos diferentes recibe el nombre de Epistasia. Se dice que un gen o grupo de genes epistásicos suprimirá la expresión de un gen o grupo de genes de un alelo diferente y que denominamos hipostáticos.
Veamos un ejemplo extremo: el síndrome del cabello acanalado o impeinable es una alteración autosómica dominante que hace que en el pelo aparezca un surco o canal longitudinal que provoca que el pelo se vuelvan muy difíciles de peinar debido a su disposición. Si un individuo con esta alteración tuviese una alopecia congénita hereditaria, el pelo acanalado no sería visible (ya que el individuo, sencillamente, carecería de pelo). Pero, al poseer el genotipo del pelo acanalado, podría transmitirlo a su descendencia. Es decir, aunque es un alelo dominante, no aparece porque otro gen lo oculta. Pero puede ser transmitido a su descendencia, pudiendo aparecer en sus descendientes siempre y cuando estos no expresasen también los genes relativos a la alopecia congénita.
Imaginemos otro ejemplo. En los hámster, un determinado gen que denominamos A hace que el ratón presente el pelo marrón o agoutí y que es dominante sobre un determinado gen que denominamos a y que hace que el ratón tenga pelo negro. Si cruzamos dos líneas híbridas Aa, sería esperable que 3/4 partes de la descendencia presentasen el pelo marrón, mientras que 1/4 parte tendrían el pelo negro. Pero si ambas líneas son también heterocigóticas para el albinismo, que hace que el pelo aparezca de color blanco (el albinismo se debe a un gen autonómico recesivo), las proporciones cambiarán, ya que los individuos albinos no tendrán coloración en el pelo (ni negro, ni marrón), por lo que las proporciones de la descendencia será 9/16 marrón, 3/16 negro y 4/16 blanco (o albino).
Debemos darnos cuenta que la proporción entre marrón y negro pasa de 4 a 1 si no apareciese interferencia con el genotipo albino, frente a la proporción 3 a 1 que encontramos cuando se nos cruza el genotipo albino.
Además, como vemos, parte de los individuos albinos transmitirán a la descendencia el genotipo marrón y parte el genotipo negro, sin que podamos apreciar ninguno de los dos genotipos.
Herencia Poligénica.
Algunas características fenotípicas no son controlados por un solo gen, sino por un grupo de genes. Existen muchos ejemplos, siendo la altura o el color de la piel o el color de ojos tres ejemplos clásicos en seres humanos.
Al existir varios genes para controlar un solo carácter, existen muchas herencias intermedias. Por este motivo, no podemos calificar a las personas sencillamente como altas o bajas, ni podemos decir que una persona es sencillamente morena o rubia, existiendo un amplio grado de niveles intermedios.
Cuando un carácter se asocia a una herencia poligñenica, la distribución de los genotipos dentro de las poblaciones suele responder a gráficas en forma de campana, con una serie de fenotipos que tenderán a ser más abundantes y que corresponderán a aquellos que se caracterizan por alelos dominantes en situación de heterocigosis, frente a otros que aparecerán con menos frecuencia de forma gradual y que se caracterizarán por alelos recesivos o por los grupos de individuos con todos los genotipos en homocigosis.
Es decir, los genotipos con mayores proporciones de homocigosis tenderán a ser más infrecuentes, frente a los heterocigóticos que además manifestarán principalmente a los alelos dominantes.
Los cilios y flagelos son apéndices móviles con dos funciones fundamentales: provocar movimiento y desplazamiento de la célula, como ocurre por ejemplo con los espermatozoides o los flagelos algunos protozoos, o para movilizar el medio externo, como ocurre de las células del epitelio respiratorio, que usan sus cilios para promover el movimiento de la mucosidad.
Cilios y flagelos poseen una estructura interna común. Los flagelos de procariotas son estructuras muy simples constituidas por proteínas fibrilares que componen el cuerpo del cilio, sin recubrimiento membranas o y que tienen el mecanismo que origina el movimiento en el interior de la célula.
Los cilios y flagelos eucariotas son mucho más complejos y se basan en una estructura constituida por microtúbulos dispuestos de forma muy ordenada. Están recubiertos por membrana y el movimiento viene originado por desplazamientos de los microtúbulos del interior del apéndice.
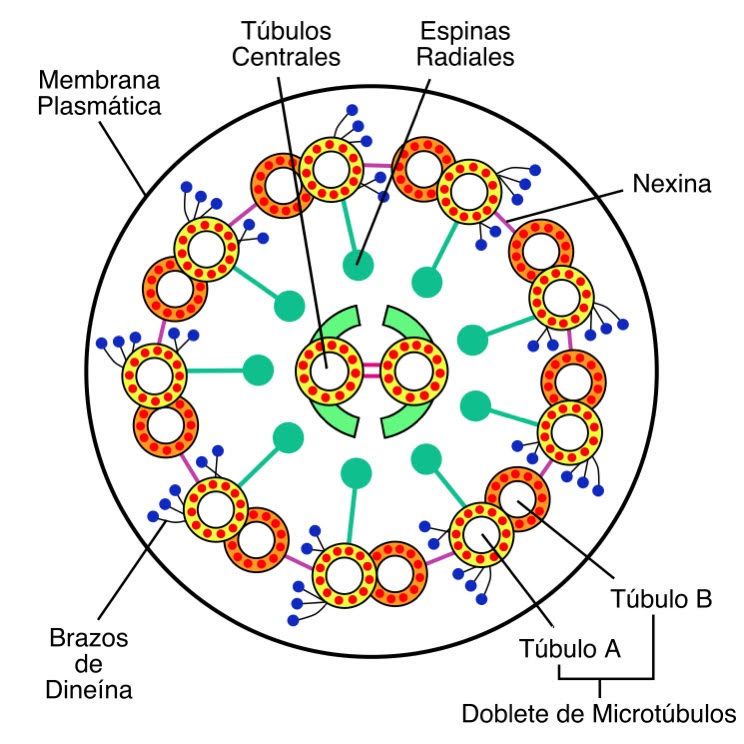
Los microtúbulos dentro del apéndice constituyen una conformación denominada Axonema y se organizan en una estructura 9+2, con nueve microtúbulos formando una estructura de doble anillo en la parte periférica, y dos tubos centrales.
Los dobletes externos llevan ancladas dos moléculas de una proteína que promueve el desplazamiento de unos microtúbuos respecto a otros y que se denomina Dineína. Es decir, la dineina conecta un doblete con el doblete vecino y es la encargada de que los microtúbulos se desplacen entre sí, mediante consumo de ATP, originando así el movimiento del cilio o flagelo. Este movimiento, por lo tanto, parte del interior mismo del apéndice.
La estructura de los deobletes, además,se estabiliza mediante proteínas que unen a los dobletes entre sí y que se denominan Nexina.
El par de microtúbulos centrales constituyen el eje del apéndice y se unen entre sí mediante un Puente Central y a los dobletes externos mediante proteínas denominadas Espinas Radiales.
En la base del cilio o flagelo encontramos siempre una estructura electrondensa, de naturaleza fibrilar y que se denomina Cuerpo Basal. El cuerpo basal es, morfológicamente, idéntico a un centriolo y su función es controlar la disposición de los microtúbulos que constituyen el axonema del cilio o flagelo.
El movimiento de cilios y flagelos es ligermente diferente. Mientras que los flagelos tienden a realizar movimientos helicoidales que permiten el movimiento de la célula en una dirección concreta, los flagelos tienden a realizar movimientos hacia atrás y adelante, que son especialmente eficaces a la hora de mover el medio externo (como el caso de la mucosidad en las células epiteliales del tubo respiratorio).
Los glúcidos, también llamados carbohidratos, hidratos de carbono o sacáridos, son moléculas orgánicas de entre tres y siete átomos de carbono, caracterizadas por la presencia de varios grupos alcohol (polialcoholes) y un grupo aldehido o un grupo cetona (carbonilo) en su molécula.
Como veremos, son las sustancias energéticas más importantes para las células, siendo el principal combustible celular. Constituyen reservas de energía tanto en animales como en vegetales. Algunos polisacáridos, además, poseen una función estructural importante y forman parte de moléculas tan variadas como la celulosa de las paredes celulares en vegetales o de los ácidos nucleicos. Y las grandes cadenas ramificadas de azúcares del exterior de la membrana celular son usadas como sistema de reconocimiento entre distintas células.
Si bien pueden ser clasificados atendiendo a criterios funcionales, lo más habitual es hacerlo en función a su estructura química y tamaño.
Así, hablamos de glúcidos simples o monosacáridos cuando encontramos a las moléculas glucídicas aisladas, mientras que hablamos de disacáridos si dos glúcidos simples se unen entre sí y polisacáridos cuando tres o más glúcidos simples se encuentran unidas entre sí, formando cadenas de entre que a su vez pueden ser lineales o ramificadas.
En términos generales, se habla de osas para referirse a los monosacáridos y de ósidos para referirse a disacáridos y polisacáridos. Los ósidos, a su vez, pueden dividirse en dos grandes grupos: holósidos, si están compuestos solo por glúcidos y heterósidos si, además de glúcidos, podemos encontrar otros compuestos orgánicos en la molécula.
Los holósidos, a su vez, pueden clasificarse como homopolisacáridos si están formados solo por un tipo de glúcidos y heteropolisacáridos si están compuestos por distintos tipos de glúcidos.
La maltosa, compuesta por dos moléculas de glucosa, es un ejemplo de homopolisacárido, mientras que la lactosa, compuesta por una molécula de glucosa y una de galactosa es un ejemplo de hereropolisacárido.